昆虫の飛行

昆虫は、羽と飛翔能力を進化させた唯一の無脊椎動物群です。昆虫が初めて飛翔したのは石炭紀、つまり約3億年から3億5000万年前で、飛翔能力を進化させた最初の動物です。羽は、既存の四肢の側面にある付属肢から進化したと考えられています。これらの付属肢には、既に神経、関節、筋肉が備わっており、それらは他の用途に使用されていました。これらの付属肢は、当初は水上を航行するため、あるいは滑空時の降下速度を遅くするために使用されていた可能性があります。
トンボとカゲロウの2つの昆虫グループは、飛翔筋が羽に直接付着しています。他の有翅昆虫では、飛翔筋は胸部に付着しており、胸部を振動させることで羽ばたきを促します。これらの昆虫の中には、ハエや一部の甲虫など、胸部が神経インパルスの速度よりも速く振動する「非同期」神経系の進化によって、非常に高い羽ばたき頻度を達成している種もいます。
すべての昆虫が飛翔能力を持っているわけではない。翅のない昆虫の中には、進化の過程で二次的に翅を失ったものも少なくない。一方、シミのようなより原始的な昆虫は、翅を進化させなかった。アリやシロアリのような一部の真社会性昆虫では、有翅生殖階級のみが交尾期に翅を発達させ、交尾後に脱翅するのに対し、他の階級の個体は生涯翅を持たない。
非常に小さな昆虫の中には、定常空気力学ではなく、ワイス・フォッホのクラップ・アンド・フリング機構を利用して、羽の摩耗を犠牲にして大きな揚力を発生させるものがあります。多くの昆虫はホバリングして高度を維持し、姿勢を制御することができます。蛾などの一部の昆虫は、前翅と後翅が連動して機能します。
メカニズム
直行便
他の昆虫とは異なり、カゲロウ類(カゲロウ)とトンボ類(トンボとイトトンボ)の羽の筋肉は羽根の基部に直接挿入されており、羽根の基部は蝶番で連結されているため、羽根の基部を少し下方に動かすと、まるで空中でボートを漕ぐように羽根自体が持ち上がります。トンボとイトトンボは前羽と後羽の形状と大きさが似ています。それぞれが独立して機能するため、他の飛翔昆虫には見られない、方向転換や速度変更の際の微細な制御と機動性が得られます。トンボ類はすべて空中捕食者であり、常に他の空中昆虫を捕食してきました。[1]
- 直進飛行:翼に付着した筋肉。大型昆虫のみ
- トンボのスローモーション飛行


間接飛行
直接飛翔筋を持つ2つの目を除き、現生の有翅昆虫はすべて、間接飛翔筋を用いた異なる機構を用いて飛翔する。この機構は一度進化し、新翅目下綱( Neoptera )を特徴づける(共形質進化) 。これは、おそらく偶然ではないが、新翅目昆虫が静止時に翅を腹部に折り畳むことを可能にする翅折り畳み機構の出現と一致する(ただし、この能力は蝶など一部のグループでは二次的に失われている)。[1]
しかし、すべての新翅目昆虫に共通するのは、胸部の筋肉の働きである。これらの筋肉は翅に付着するのではなく、胸部に付着してそれを変形させる。翅は胸郭外骨格の延長であるため、胸部の変形は翅の動きにも影響を与える。背部に沿った一連の縦走筋が胸部を前後に圧迫し、胸部背面(胸背板)を上方に反らせ、翅を下方に折り曲げる。背板から胸骨にかけての別の一連の筋が胸背板を再び下方に引っ張り、翅を上方に折り曲げる。[1] [2]
- 間接飛行:胸郭の筋肉が振動する。a
:翼、
b:主・二次飛行関節、
c:背腹飛行筋、
d:縦走筋


1秒間に100回未満しか羽ばたかない昆虫は、同期筋を用いています。同期筋とは、神経インパルスごとに1回収縮するタイプの筋肉です。同期筋は一般的に非同期筋よりも出力が低く、効率も低いため、複数の異なる昆虫系統において非同期飛翔筋が独立して進化してきたと考えられます。[3]
マルハナバチのように羽をより速く羽ばたかせる昆虫は非同期筋を使う。非同期筋とは、神経インパルスごとに複数回収縮するタイプの筋肉である。これは、筋肉の緊張が解放されることで筋肉が再び収縮するように刺激されることによって実現され、これは単純な神経刺激だけよりも速く起こり得る。[4]これにより、羽ばたきの頻度が神経系がインパルスを送ることができる速度を超えることが可能となる。非同期筋は、高等な新翅目(甲虫目、双翅目、膜翅目)の一部に現れた最終的な改良点の1つである。全体的な効果として、多くの高等な新翅目は、直接飛行する筋肉を持つ昆虫よりもはるかに速く羽ばたくことができる。[1]
非同期筋は、定義上、神経系による比較的粗い制御下にあります。この進化的トレードオフのバランスを取るため、間接飛行を進化させた昆虫は、翼のストロークを細かく制御するための独立した神経筋系も発達させました。[5]「直接筋」として知られるこれらの筋肉は、翼のヒンジを構成する小板に直接付着し、運動ニューロンからの1:1のインパルスによって収縮します。[6]最近の研究では、翼のヒンジにおける複雑な非線形筋力学とそれが翼端軌道に与える影響について検討が始まっています。[7]
空気力学
昆虫の飛行には2つの基本的な空気力学的モデルがあります。前縁渦を発生させるモデルと、クラップアンドフリングを使用するモデルです。[8] [9]
前縁渦
ほとんどの昆虫は、螺旋状の前縁 渦を作り出す方法を用いる。これらの羽ばたきは、2つの基本的な半ストロークで移動する。下降ストロークは上方から後方に始まり、下方から前方に急降下する。次に、翼は素早く反転(回外)し、前縁が後方を向く。次に、上昇ストロークで翼は上方から後方に押し出される。その後、翼は再び反転(回内)し、別の下降ストロークが発生する。同期飛翔筋を持つ昆虫の 周波数範囲は通常、5~200ヘルツ(Hz)である。非同期飛翔筋を持つ昆虫では、羽ばたき周波数は1000 Hzを超えることがある。昆虫がホバリングしているとき、2つのストロークにかかる時間は同じである。しかし、より遅い下降ストロークが推力を生み出す。[8] [9]
昆虫の飛行を理解するには、主要な力の特定が不可欠です。羽ばたき翼を理解するための最初の試みは、準定常状態を仮定していました。これは、任意の時点における翼上の気流が、同じ迎え角で羽ばたかない定常状態の翼上の流れと同じであると仮定したことを意味します。羽ばたき翼を多数の静止位置に分割し、それぞれの位置を分析することで、翼に瞬間的に作用する力のタイムラインを作成することが可能になります。計算された揚力は3分の1ほど小さすぎることが判明したため、研究者たちは空気力を提供する非定常現象が存在するに違いないと考えました。羽ばたき翼付近の流れを近似しようと試みる解析モデルがいくつか開発されていました。一部の研究者は、回外時に力のピークが生じると予測しました。ショウジョウバエの動的スケールモデルを用いて、これらの予測された力は後に確認されました。他の研究者は、回外および回内時の力のピークは、並進現象とは根本的に異なる未知の回転効果によって引き起こされると主張しました。この主張には異論もある。数値流体力学を用いて、回転効果は存在しないと主張する研究者もいる。彼らは、大きな力は前のストロークによって生じた後流との相互作用によって引き起こされると主張している。[8] [9]
上述の回転効果と同様に、羽ばたきに伴う現象も完全には理解されておらず、また合意に達していません。すべてのモデルが近似値であるため、モデルによって無視できると考えられる効果が除外されています。たとえば、1925 年にハーバート A. ワグナーが提唱したワグナー効果[10]によれば、傾斜した翼が静止状態から加速されると、粘性により循環がゆっくりと定常状態まで上昇します。この現象は、予測よりも小さい揚力値を説明するものです。通常、追加された揚力の発生源を見つけることが問題となっています。昆虫の飛行に典型的なレイノルズ数の流れでは、この効果は無視できると主張されてきました。レイノルズ数は乱流の尺度であり、レイノルズ数が低い場合は流れは層流(滑らか) になり、レイノルズ数が高い場合は乱流になります[11] 。ワグナー効果は、少なくとも 1 つのモデルでは意識的に無視されていました。[9]昆虫の飛行中に起こる最も重要な現象の一つは、前縁吸引である。この力は効率の計算に重要である。前縁吸引の概念は、鋭利な縁を持つデルタ翼の渦揚力を説明するために、1988年にDG EllisとJL Stolleryによって初めて提唱された。[12]迎え角が大きい場合、流れは前縁上で剥離するが、後縁に達する前に再付着する。この剥離した流れの泡の中に渦が存在する。迎え角が非常に大きいため、多くの運動量が下向きの流れに伝達される。これら2つの特徴により、大きな揚力といくらかの追加の抗力が生じる。しかし、重要なのは揚力である。流れが剥離しているにもかかわらず、依然として大きな揚力を生み出すため、この現象は失速遅延と呼ばれ、1945年にH.ヒンメルスカンプによって航空機のプロペラで初めて観察されました。[13]この効果は昆虫の羽ばたき飛行において観察され、準定常モデルの不足を補うのに十分な揚力を生み出すことが証明されました。この効果は、カヌーの漕ぎ手がスカリングのドローストロークで利用しています。[8] [9]
羽ばたき翼に及ぼす影響はすべて、空気力学的現象の3つの主要な発生源、すなわち前縁渦、翼にかかる定常空気力、そして前回のストロークによる翼の後流との接触に還元できます。飛翔昆虫の大きさは約20マイクログラムから約3グラムです。昆虫の体重が増加すると、翼面積が増加し、羽ばたき周波数は低下します。大型昆虫では、レイノルズ数(Re)は10000にも達し、流れが乱流になり始めます。小型昆虫では、レイノルズ数は10程度にまで低下します。これは、小型昆虫にとって粘性効果がはるかに重要であることを意味します。[9] [14] [15]
昆虫の飛行におけるもう一つの興味深い特徴は、体の傾きです。飛行速度が増加すると、昆虫の体は機首を下げ、より水平に傾く傾向があります。これにより前面面積が減少し、ひいては体の抗力も減少します。前進速度が増加すると抗力も増加するため、昆虫は飛行効率の向上が求められるにつれて、より効率的な飛行を実現しています。さらに、下降運動における幾何学的な迎え角を変化させることで、昆虫は可能な限り多くの運動において最適な飛行効率を維持することができます。一般的な推力の発生は揚力に比べて比較的小さいです。揚力は昆虫の体重の3倍以上になることもありますが、最高速度時でも推力は体重の20%程度にしか達しません。この力は、主に羽ばたき運動におけるより弱い上昇運動によって発生します。[9] [16]
拍手と投げ飛ばし
デンマークの動物学者トルケル・ヴァイス=フォグによって発見されたクラップ・アンド・フリング、またはヴァイス=フォグ機構は、小型昆虫の飛行中に利用される揚力発生方法です。[17]昆虫のサイズが1 mm未満になると、粘性力が支配的になり、翼型による揚力発生の効率が大幅に低下します。クラップ位置から、2枚の翼は離れ、後縁を中心に回転します。その後、翼は分離し、下降ストロークの終わりまで水平方向にスイープします。次に、翼は回内し、上昇ストロークの漕ぎ動作中に前縁を利用します。クラップ動作が始まると、前縁は接触し、隙間がなくなるまで一緒に回転します。当初は翼が接触していると考えられていましたが、いくつかの事例から翼の間に隙間があることが示され、それが空気力学的利点をもたらしていることが示唆されています。[18] [19]
クラップアンドフリング機構による揚力生成は、運動中のいくつかの過程で起こる。まず、単一の翼運動では十分な揚力を生成できないため、この機構は翼間相互作用に依存している。[20] [21] [22] フリング運動で翼が後縁の周りを回転すると、できた隙間に空気が流れ込み、強い前縁渦が発生し、2 番目の渦が翼端で発生する。3 番目のより弱い渦が後縁で発生する。[19]発生する渦の強さは、フリング運動開始時の翼間分離の初期の隙間に一部依存する。翼間隙間が減少すると、抗力が大きくなるが、揚力は大きくなる。フリング中のヒービング運動の実施、[23]柔軟な翼、[21]および遅延失速機構が、渦の安定性と付着を強化することがわかった。[24]最後に、低レイノルズ数飛行(層流)中の全体的な揚力生成の低下を補うために、小さな昆虫は、より大きな昆虫に匹敵する翼端速度を生成するために、より高いストローク周波数を持つことが多い。[24]
予想される全体的な抗力は、翼が分離して回転する必要がある背部の羽ばたき運動中に最大になります。[22]大きな抗力の減衰は、いくつかのメカニズムによって発生します。柔軟な翼は、剛翼と比較して、羽ばたき運動における抗力を最大50%削減し、さらに翼のストローク全体を通して全体的な抗力を削減することがわかっています。[21] Encarsia formosaに見られるように、翼端の剛毛は流れに多孔性を引き起こし、抗力を増強および削減しますが、揚力生成は低くなります。[25]さらに、羽ばたき前の翼間の分離は、全体的な抗力の影響に重要な役割を果たします。翼間の距離が増加すると、全体的な抗力は減少します。[18]
拍手と投げ飛ばしのメカニズムは、海洋軟体動物の Limacina helicina(海の蝶)にも採用されています。[26] [27]野菜の葉の鉱夫Liriomyza sativae(ハエ)などの一部の昆虫は、部分的な拍手と投げ飛ばしを利用して、翼の外側部分のみにこのメカニズムを適用し、ホバリング時に約7%の揚力を増加させます。[24]
- Sane 2003 に基づくクラップ アンド フリング飛行機構
- 拍手1: 背中の上で翼を閉じる
- 拍手2:前縁が接触し、翼が前縁を中心に回転し、渦が形成される
- 3回目の拍手:後縁が閉じ、渦が消え、翼が閉じて推進力を与える



- 黒(曲線)矢印:流れ;青矢印:誘導速度;オレンジ矢印:翼にかかる正味の力
- 投げ飛ばし1:翼が後縁を中心に回転して飛び散る
- フリング2:前縁が離れ、空気が流れ込み、揚力が増加する
- フリング3:前縁に新しい渦が形成され、後縁の渦は互いに打ち消し合い、流れが速まる可能性がある(Weis-Fogh 1973)



支配方程式
流体中を移動する翼は、空気力学の慣例に従って流体力を受ける。翼に対して流れの方向に垂直な力の成分は揚力(L)と呼ばれ、流れの反対方向の力の成分は抗力(D)と呼ばれる。ここで考察するレイノルズ数において、適切な力の単位は1/2(ρU 2 S)である。ここで、ρは流体の密度、Sは翼面積、Uは翼速である。無次元の力は揚力係数(C L)と抗力係数(C D)と呼ばれ、それぞれ以下の式で表される:[8]

C LとC D は、流れが定常である場合にのみ定数となる。翼のような特殊な物体は、小さな迎え角で流体を横切る際に定常状態に達することがある。この場合、翼周りの非粘性流れは、非貫通境界条件を満たすポテンシャル流で近似することができる。2次元翼のクッタ・ジューコフスキー定理はさらに、流れが鋭い後縁から滑らかに離れることを仮定しており、これが翼周りの全循環を決定する。対応する揚力はベルヌーイの定理(ブラシウスの定理)によって与えられる。[8]

鳥や昆虫の周りの流れは非圧縮性であるとみなすことができます。マッハ数、つまり空気中の音速に対する相対速度は典型的には1/300で、翼の振動数は約10~103Hzです。支配方程式として、滑りなし境界条件に従うナビエ・ストークス方程式を用いると、次式が得られます。 [8]

ここで、u (x, t) は流れ場、p は圧力、ρ は流体の密度、ν は動粘性、u bd は境界における速度、u s は固体の速度です。長さスケール L と速度スケール U を選択すると、方程式はレイノルズ数 R e =uL/ν を含む無次元形式で表すことができます。昆虫の羽と翼型には、2つの明らかな違いがあります。昆虫の羽ははるかに小さく、羽ばたきます。トンボを例に挙げると、翼弦長 (c) は約1 cm (0.39 インチ)、翼長 (l) は約4 cm (1.6 インチ)、翼周波数 (f) は約40 Hzです。翼端速度 (u) は約 1 m/s (3.3 ft/s) で、対応するレイノルズ数は約 103 です。小さいほうの典型的なカルシドイドバチの羽根の長さは約 0.5~0.7 mm (0.020~0.028 インチ) で、羽根の羽ばたき周波数は約 400 Hz です。レイノルズ数は約 25 です。昆虫の飛行時のレイノルズ数の範囲は約 10 ~ 10 4で、理論上都合の良い 2 つの限界値、つまり翼の周りの非粘性の定常流と遊泳細菌が経験するストークス流の間にあります。このため、この中間範囲は十分に理解されていません。一方、これは私たちが目にする物の間でおそらく最も普遍的な領域です。落ち葉や種子、魚、鳥はすべて、昆虫の周りに見られるのと同様の非定常流に遭遇します。[8]弦方向レイノルズ数は次のように記述される。

そして


ここで、 は平均翼弦長、は翼端の速度、はストローク振幅、はビート周波数、は回転半径、は翼面積、 は翼端を含む翼の長さです。







レイノルズ数に加えて、少なくとも2つの関連する無次元パラメータがあります。翼には3つの速度スケールがあります。すなわち、翼体に対する羽ばたき速度(u)、翼体の前進速度(U 0)、そしてピッチング速度(Ω c)です。これらの比は、U 0 / uとΩc/ uという2つの無次元変数を形成します。前者はしばしば前進比と呼ばれ、縮約周波数fc / U 0とも関連しています。[8]
昆虫の羽が剛体である場合(例えばショウジョウバエの羽はほぼ剛体である)、固定物体に対する羽の相対運動は3つの変数、すなわち球座標における羽根先の位置(Θ(t),Φ(t))、および根元と羽根先を結ぶ軸の周りのピッチング角(ψ(t))で記述できる。羽根要素解析に基づいて空気力を推定するには、迎え角(α)を決定することも必要である。ホバリングする昆虫では、翼幅の70%における典型的な迎え角は25°から45°の範囲である(ハチドリでは15°)。多くの昆虫について豊富なデータが得られるにもかかわらず、ストローク中のαの時間変化を報告している実験は比較的少ない。これらの例としては、繋留されたイナゴとハエの風洞実験、およびショウジョウバエの自由ホバリング飛行が挙げられる。[8]
翼端軌道は比較的測定が容易であるため、より頻繁に報告されています。例えば、重量を支えるのに十分な揚力を生み出す飛行シーケンスのみを選択すると、翼端が楕円形を描いていることがわかります。他の昆虫では、交差しない形状も報告されています。正確な形状に関わらず、翼端が下方に押し下げられる動きは、昆虫が重量を支えるために揚力に加えて空気抵抗を利用している可能性を示唆しています。[8]
ホバリング
| 飛行パラメータ[28] | 速度(m/s) | ビート/秒 |
|---|---|---|
| エスニッドトンボ | 7.0 | 38 |
| ホーネット | 5.7 | 100 |
| ハチドリガ | 5.0 | 85 |
| アブ | 3.9 | 96 |
| ハナアブ | 3.5 | 120 |
| バンブルビー | 2.9 | 130 |
| ミツバチ | 2.5 | 250 |
| イエバエ | 2.0 | 190 |
| イトトンボ | 1.5 | 16 |
| サソリバエ | 0.49 | 28 |
| オオモンシロチョウ[29] | 2.5 | 12 |
| アザミウマ(パタパタと飛ばす)[24] | 0.3 | 254 |
.jpg/1280px-Hoverfly_September_2007-7_(cropped).jpg)
多くの昆虫は、羽を素早く羽ばたかせることで空中で一点に留まる、つまりホバリングすることができます。そのためには、横方向の安定性と揚力の発生が必要です。揚力は主に下降運動によって生じます。羽が周囲の空気を押し下げると、空気の反作用によって昆虫は上方に押し上げられます。ほとんどの昆虫の羽は、上昇運動中に羽にかかる力が小さくなるように進化しています。下降運動と下降運動によって昆虫はそれぞれ上方と下方に押し上げられるため、昆虫は振動し、最終的に同じ位置に留まります。[14]
昆虫が羽ばたく間に落下する距離は、羽ばたく速さに依存する。羽ばたきが遅いほど、落下間隔は長くなり、羽ばたき間の落下距離は長くなる。昆虫が一定の振幅を安定させるために必要な羽ばたき周波数を計算することができる。計算を簡略化するために、羽が下降している間は揚力は有限の一定値であり、上昇している間は揚力はゼロであると仮定する必要がある。上昇する羽ばたきの時間間隔Δtの間に、昆虫は重力の影響を受けてhの距離を落下する。[14]

上向きのストロークによって昆虫は元の位置に戻ります。通常、昆虫の垂直位置の変化は0.1mm(つまりh = 0.1mm)以下である必要があります。この場合、自由落下の最大許容時間は[14]です。

翼の上下運動の持続時間はほぼ同じであるため、完全な上下運動の周期TはΔrの2倍である。つまり、[14]

羽ばたきの周波数fは1秒あたりの羽ばたきの回数を意味し、次の式で表される:[14]

例として用いた振動数は110回/秒であり、これは昆虫に見られる典型的な振動数である。蝶の振動数は10回/秒程度とはるかに低く、ホバリングはできない。他の昆虫は1000回/秒の振動数を出すことができるかもしれない。昆虫を元の垂直姿勢に戻すには、下降運動中の平均上向きの力F avが昆虫の体重の2倍に等しくなければならない。昆虫の体にかかる上向きの力は半分の時間しかかからないため、昆虫にかかる平均上向きの力は単に体重と同じであることに注意されたい。[14]
電源入力
ここで、質量m = 0.1gの昆虫を例にとると、ホバリングを維持するために必要な力を計算することができます。この場合、下降時に両羽根が加える平均力F avは、重量の2倍になります。羽根にかかる圧力は羽根全体にわたって均一に分散されるため、各羽根が生み出す力は、羽根の中央部の一点に作用すると想定できます。下降時に、羽根の中心は垂直距離dを移動します。[14]昆虫が各下降時に行う全仕事は、力と距離の積です。つまり、

羽根が70°の角度で羽根を振るとすると、羽根の長さが1cmの昆虫の場合、dは0.57cmとなる。したがって、両羽根が1回の羽根振りで行う仕事は以下の通りである。[14]

このエネルギーは昆虫を重力に逆らって持ち上げるために使用される。昆虫の質量を1回の下降ストロークで0.1mm持ち上げるのに必要なエネルギーEは[14]である。

これは消費されるエネルギー全体から見ればごくわずかな割合であり、明らかにエネルギーの大部分は他のプロセスで消費されています。この問題をより詳細に分析すると、羽根によって行われた仕事は主に空気の運動エネルギーに変換され、この運動エネルギーは羽根の下向きのストロークによって加速されることが分かります。出力とは1秒間に行われる仕事量です。例として挙げた昆虫は1秒間に110回の下向きのストロークを行います。したがって、その出力Pは1秒間のストローク数であり、つまり出力Pは[14]となります。

出力
ホバリングに要する力の計算において、用いられた例では動いている羽の運動エネルギーは考慮されていませんでした。昆虫の羽は軽いとはいえ、有限の質量を持ちます。そのため、羽は移動する際に運動エネルギーを持ちます。羽は回転運動をするため、1回の羽ばたきにおける最大運動エネルギーは次のようになります。[14]

ここで、Iは翼の慣性モーメント、 ωmaxは翼のストローク中の最大角速度である。翼の慣性モーメントを求めるために、翼は一端を軸とした細い棒で近似できると仮定する。すると、翼の慣性モーメントは以下のようになる:[14]

ここで、 lは翼の長さ(1 cm)、mは2枚の翼の質量(典型的には10 −3 g)である。最大角速度ωmaxは、翼中心における最大線速度νmaxから計算できる。 [14]

各ストロークの間、翼の中心は平均線速度νavで移動する。νavは、翼の中心が移動した距離dをストロークの持続時間Δtで割った値で与えられる。前述の例では、d = 0.57 cm、Δt = 4.5×10 −3 sである。したがって、次の式が成り立つ。 [14]

翼の速度は、翼のストロークの開始時と終了時の両方でゼロです。つまり、最大線速度は平均速度よりも高くなります。速度が翼の軌跡に沿って(正弦波状に)振動すると仮定すると、最大速度は平均速度の2倍になります。したがって、最大角速度は次のようになります。[14]

したがって運動エネルギーは次のようになる:[14]

翼運動の各サイクルには2回の翼のストローク(上昇ストロークと下降ストローク)があるため、運動エネルギーは2×43 = 86 エルグとなります。これは、ホバリング自体に消費されるエネルギーとほぼ同じです。[14]
弾性

昆虫は、羽が加速するときに筋肉から供給される運動エネルギーを得る。羽がストロークの終わりに向かって減速し始めると、このエネルギーは消散しなければならない。下降ストロークの間、運動エネルギーは筋肉自体によって消散され、熱に変換される(この熱は体幹温度を維持するために使用されることがある)。一部の昆虫は、羽の上向きの動きで運動エネルギーを利用して飛行を補助することができる。これらの昆虫の羽の関節には、レジリンと呼ばれる弾性のあるゴムのようなタンパク質のパッドが含まれている。羽が上昇する間、レジリンは引き伸ばされる。羽の運動エネルギーは、伸びたレジリンの位置エネルギーに変換され、バネのようにエネルギーを蓄える。羽が下降すると、このエネルギーは放出され、下降ストロークを補助する。[14]
いくつかの単純化した仮定を用いることで、伸張したレジリンに蓄えられるエネルギー量を計算できる。レジリンは複雑な形状に曲げられているが、この例では面積Aと長さAの直線棒として計算している。さらに、伸張中はレジリンはフックの法則に従うと仮定する。しかし、これは厳密には正しくない。レジリンはかなりの量に伸張するため、伸張の過程で面積とヤング率の両方が変化するからである。伸張したレジリンに蓄えられる位置エネルギーUは、以下の式で表される。[14]

ここでEはレジリンのヤング率であり、1.8×10 7 dyn /cm 2と測定されている。典型的には、ハチほどの大きさの昆虫では、レジリンの体積は長さ2×10 −2 cm、面積4×10 −4 cm 2の円筒に相当する。この例では、レジリン棒の長さは伸長すると50%増加する。つまり、Δℓは10 −2 cmである。したがって、この場合、各羽のレジリンに蓄えられる位置エネルギーは、以下の式で表される:[14]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
ハチサイズの昆虫の両羽に蓄えられるエネルギーは36エルグで、これは羽の跳ね上がり運動エネルギーに匹敵します。実験では、羽の運動エネルギーの最大80%がレジリンに蓄えられていることが示されています。[14]
翼カップリング
{kind=link}

鱗翅目昆虫などの一部の四翅昆虫目は、成虫において形態学的な翅連結機構を発達させ、機能的に二翅昆虫となっている。[30]最も原始的な形態を除くすべての昆虫がこの翅連結構造を示す。[31]
そのメカニズムには、頬骨型、小帯網膜型、および膨大型の3つのタイプがあります。[32]
- より原始的なグループでは、前翅の基部、すなわち後縁基部付近に拡大した葉状の領域があり、飛行中は後翅の下に折りたたまれる羽片。 [31] [33]
- 他のグループでは、後翅に小帯があり、前翅の支帯の下に引っ掛かる。[33]
- ほとんど全ての蝶類とカイコ上科(スズメガ科を除く)では、羽を連結する小帯と支帯の配置は見られない。代わりに、後羽の上腕部の拡大部分が前羽に広く重なり合っている。特定の機械的な接続がないにもかかわらず、両羽は重なり合い、同調して動作する。前羽の力強い動きが後羽を同期して押し下げる。このタイプの連結は小帯型連結のバリエーションであるが、小帯と支帯は完全に失われている。[30] [34]
生化学
昆虫の飛翔に関する生化学は、これまで多くの研究の対象となってきました。多くの昆虫は飛翔のエネルギー源として炭水化物と脂質を利用しますが、多くの甲虫やハエはアミノ酸である プロリンをエネルギー源としています。[35]また、一部の種は複数のエネルギー源を組み合わせて利用しており、 Manduca sextaなどの蛾は、飛翔前のウォーミングアップに炭水化物を利用しています。[36]
感覚フィードバック
昆虫は感覚フィードバックを利用して飛行を維持し、制御します。昆虫は触角[37] 、棍棒[38] 、羽[39]からの情報を用いて飛行姿勢、羽ばたきの振幅、羽ばたきの頻度を制御します。
進化と適応
{kind=link}
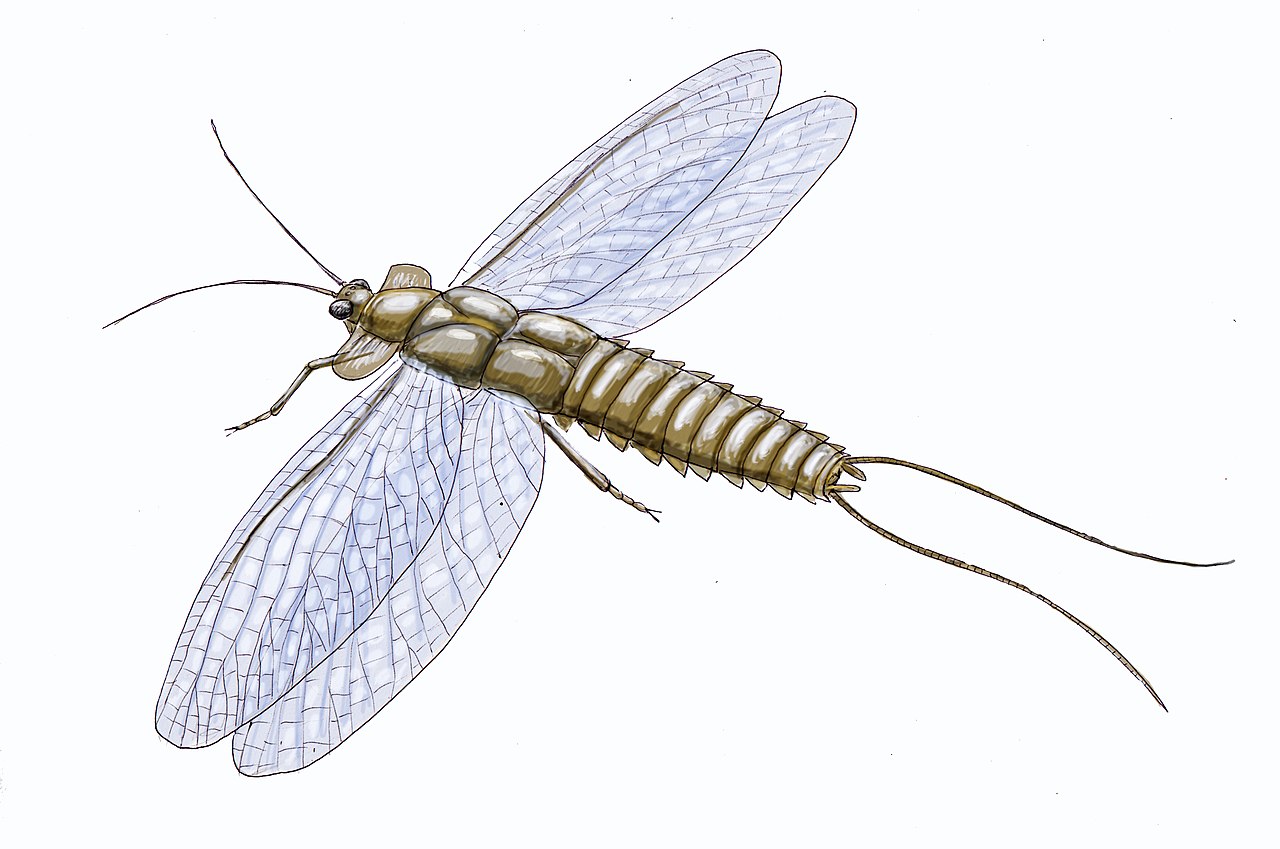
約3億5000万年から3億年前の石炭紀に、まだ2つの大きな陸地しかなかった頃に、昆虫が飛び始めました。[40]最も古い有翅昆虫の化石の一つに、石炭紀前期の古翅目昆虫であるデリッツチャラがあります。 [41]リニオグナタ類はそれより古く、デボン紀前期のものですが、それが有翅昆虫であったのか、あるいは昆虫であったのかは定かではありません。[42] [43]
昆虫の羽がなぜ、どのように発達したかはよくわかっていません。これは主に、下部石炭紀の発達期に関する適切な化石が乏しいためです。昆虫の飛翔の起源については、歴史的に主に 3 つの学説がありました。1 つ目は、カゲロウの水生幼虫に見られる可動式の腹部の 鰓が変形したものであるというものです。系統ゲノム解析により、バッタを含む有翅昆虫のグループである多翅目は陸生の祖先から進化したため、鰓から羽が進化した可能性は低いことが示唆されています。[44]カゲロウの幼虫の跳躍行動に関する追加研究により、気管鰓は昆虫の下降を導く役割を果たしていないことが判明し、この進化仮説を否定するさらなる証拠となっています。[45]これにより、歴史的には 2 つの学説が残ります。1 つは、羽が胸部背板の延長である傍胸葉から発達したもの、もう 1 つは、すでに筋肉を含んでいた脚節の変形から発生したというものです。[46]
上鰓(腹部鰓)仮説
.JPG){kind=link}
.JPG/1280px-Mayfly_nymph_dorsal_view_wing_buds_paired_gills_(rotated).JPG)
1871年のランドワ、1873年のラボック、 1877年のグレーバー、 1905年のオズボーンなど、多くの昆虫学者[47] は、昆虫の羽の起源として、カゲロウの幼虫など多くの水生昆虫に見られる可動性の腹部鰓が考えられたと示唆している。この説によれば、これらの気管鰓は呼吸器系の出口として発達し、時を経て移動手段として変化し、最終的に羽へと進化した。気管鰓には小さな翼片が備わっており、それらは絶えず振動し、独自の小さな直筋を持つ。[48]
傍脳(背部)仮説
傍胸葉あるいは背側体壁仮説は、1875年にフリッツ・ミュラー[49]により提唱され、1916年にG・クランプトン[47] 、 1978年にヤルミラ・クカロワ=ペック[50]、1981年にアレクサンダー・P・ラスニツィン[51]らにより再考され、昆虫の翅は傍胸葉から発達したと示唆している。傍胸葉は昆虫の化石に見られる前適応で、跳躍や落下時の安定性に役立ったと考えられる。この仮説を支持するのは、ほとんどの昆虫が枝登り中に驚くと地面に飛び降りて逃げる習性である。このような葉はパラシュートとして機能し、昆虫がより柔らかく着地することを可能にしたと考えられる。この理論では、これらの葉が徐々に大きくなり、後の段階で胸部と関節が発達したと示唆している。さらに後になって、これらの粗い翅を動かす筋肉が出現する。このモデルは、翼の有効性がパラシュートから始まり、滑空、そして最終的には能動飛行へと段階的に向上していくことを示唆している。しかしながら、翼の関節や筋肉の発達を示す確かな化石証拠が不足していること、そして関節や脈が一見自発的に発達したように見えることが、この理論にとって大きな障害となっている。そのため、この分野の専門家からは広く否定されている。[48]
胸膜出口仮説
{kind=link}

1990年、JWHトゥルーマンは、羽は原始的な節足動物の肢の内側と外側にある付属肢であるエンドイットとエキジットから適応したという説を提唱し、これは胸膜仮説とも呼ばれました。これは、1945年にゴールドシュミットが行ったキイロショウジョウバエ(Drosophila melanogaster)の研究に基づいています。この研究では、「ポッド」(ポドメア、肢節)と呼ばれる変異体が、正常な羽を変形させる突然変異を示しました。その結果、3関節の脚構造といくつかの付属肢が追加され、通常は翼の肋骨面がある足根が欠損していると解釈されました。この突然変異は、脚ではなく、背側のエキジットとエンドイットの融合を強く示唆する証拠として再解釈され、付属肢はこの仮説に非常によく適合しました。羽の進化に必要な神経支配、関節構造、筋肉組織は、既に肢節に存在していました。[46]
その他の仮説
他の仮説としては、ヴィンセント・ウィグルスワースが1973年に提唱した、翼は放射器として使われていた胸部の突起から発達したという説がある。[48]
エイドリアン・トーマスとオーケ・ノーバーグは2003年に、一部のカワゲラに見られるように、翼は当初水面を航行するために進化した可能性があると示唆した。[52] [53]
スティーブン・P・ヤノビアクとその同僚は2009年に、翼は有翼昆虫の無翼の姉妹種である無翼脊椎動物に見られる飛行前の現象である、指向性滑空降下から派生したものであると提唱した。[54]
二重起源
アヴェロフ[55]、丹羽[56] 、エリアス=ネト[57]をはじめとする生物学者たちは、古生物学的証拠に加えて進化発生学を用いて、昆虫の羽の起源を探り始めています。この研究は、羽が背板と胸板の両方の構造と連続的に相同性を持っていることを示唆しており、何世紀にもわたる論争に決着をつける可能性を秘めています。[58]ヤクブ・プロコプらは2017年、古生代の幼虫の羽肉から、羽が実際にそのような二重の起源を持っていたことを示す古生物学的証拠を発見しました。[59]
参照
参考文献
- ^ abcd Chapman, AD (2006). オーストラリアと世界の生物種の数. キャンベラ:オーストラリア生物資源研究. 60ページ. ISBN 978-0-642-56850-2. 2009年5月19日時点のオリジナルよりアーカイブ。2015年9月15日閲覧。
{{cite book}}: CS1 maint: publisher location (link) - ^ Smith, DS (1965). 「昆虫の飛翔筋」. Scientific American . 212 (6): 76– 88. Bibcode :1965SciAm.212f..76S. doi :10.1038/scientificamerican0665-76. PMID 14327957.
- ^ Josephson, Robert K.; Malamud, Jean G.; Stokes, Darrell R. (2001). 「甲虫の非同期飛翔筋の効率」. The Journal of Experimental Biology . 204 (23): 4125– 4139. doi :10.1242/jeb.204.23.4125. PMID 11809787.
- ^ 「昆虫学者用語集における非同期筋の定義」 ノースカロライナ州立大学昆虫学部。2011年3月21日閲覧。
- ^ Deora, Tanvi; Gundiah, Namrata; Sane, Sanjay P. (2017-04-15). 「ハエの胸郭の力学」. The Journal of Experimental Biology . 220 (8): 1382– 1395. doi : 10.1242/jeb.128363 . PMID 28424311. S2CID 207172023.
- ^ ハイデ、GG (1983)。 「双翅目における飛行制御の神経機構」 Nachtigall、Werner (編)。ビオナレポート。 Vol. 2:昆虫飛行 II。シュトゥットガルト:グスタフ・フィッシャー。33~ 52ページ 。
- ^ Melis, Johan M.; Dickinson, Michael H. (2023-06-30). 「機械学習により昆虫の翅ヒンジの制御メカニズムが解明」BioRxiv: 生物学プレプリントサーバー. doi :10.1101/2023.06.29.547116. PMC 10327165 . PMID 37425804.
- ^ abcdefghijk Wang, Z. Jane (2005). 「昆虫の飛行を解剖する」(PDF) . Annual Review of Fluid Mechanics . 37 (1): 183– 210. Bibcode :2005AnRFM..37..183W. doi :10.1146/annurev.fluid.36.050802.121940. 2014年1月16日時点のオリジナル(PDF)からアーカイブ。2015年9月15日閲覧。
- ^ abcdefg Sane, Sanjay P. (2003). 「昆虫の飛行の空気力学」(PDF) . The Journal of Experimental Biology . 206 (23): 4191– 4208. doi : 10.1242/jeb.00663 . PMID 14581590. S2CID 17453426.
- ^ ワーグナー、ヘルベルト(1925)。 「Über die Entstehung des dynamischen Auftriebes von Tragflügeln」[翼の動的揚力の起源について]。Zeitschrift für Angewandte Mathematik und Mechanik (ドイツ語)。5 (1): 17–35。ビブコード:1925ZaMM....5...17W。土井:10.1002/zamm.19250050103。
- ^ 「変遷と乱流」. プリンストン大学. 2021年7月8日閲覧。
- ^ Ellis, DG; Stollery, JL ( 1988). 「前縁渦フラップの挙動と性能」(PDF) . ICAS 1988 エルサレム. 1988 . 国際航空科学会議: 758–765 . 2021年7月8日閲覧。
- ^ Himmelskamp, H. (1945)「回転するスクリューのプロファイル調査」ゲッティンゲン大学博士論文。
- ^ abcdefghijklmnopqrstu v Davidovits, Paul (2008). 生物学と医学における物理学.アカデミック・プレス. pp. 78– 79. ISBN 978-0-12-369411-9。
- ^ Hope, Daniel K.; DeLuca, Anthony M.; O'Hara, Ryan P. (2018-01-03). 「バイオミメティック羽ばたき翼におけるレイノルズ数の影響の調査」. International Journal of Micro Air Vehicles . 10 (1): 106– 122. doi : 10.1177/1756829317745319 .
- ^ 「航跡を捉える」サイエンティフィック・アメリカン誌、1999年6月28日。
- ^ Weis-Fogh, Torkel (1973). 「ホバリング動物の飛行適応度の簡易推定、揚力生成の新しいメカニズムを含む」. Journal of Experimental Biology . 59 : 169–230 . doi :10.1242/jeb.59.1.169.
- ^ ab Santhanakrishnan, A.; Robinson, A.; Jones, S.; Low, A.; Gadi, S.; Hendrick, T.; Miller, L. (2014). 「微小昆虫の飛行における相互作用する多孔質翼による拍動と羽ばたきのメカニズム」. The Journal of Experimental Biology . 217 (Pt 21): 3898– 4709. doi : 10.1242/jeb.084897 . PMID 25189374.
- ^ ab Kolomenskiy, D; Moffatt, H.; Farge, M.; Schneider, K. (2011). 「ホバリング昆虫のclap-fling-sweep運動の2次元および3次元数値シミュレーション」Journal of Fluids and Structures . 27 (5): 784. Bibcode :2011JFS....27..784K. doi :10.1016/j.jfluidstructs.2011.05.002.
- ^ Bennett, L. (1977). 「Clap and fling aerodynamics - an experimental evaluation」. Journal of Experimental Biology . 69 : 261–272 . doi :10.1242/jeb.69.1.261.
- ^ abc Miller, L.; Peskin, S. (2009). 「小さな昆虫の飛行における柔軟な拍動と羽ばたき」. The Journal of Experimental Biology . 212 (19): 3076– 3090. doi : 10.1242/jeb.028662 . PMID 19749100. S2CID 29711043.
- ^ ab Lehmann, F.-O.; Sane, S.; Dickinson, M. (2005). 「昆虫の羽ばたきにおける翼間相互作用の空気力学的影響」. Journal of Experimental Biology . 208 (Pt 16): 3075– 3092. doi : 10.1242/jeb.01744 . PMID 16081606. S2CID 7750411.
- ^ Lehmann, F.-O.; Pick, S. (2007). 「羽ばたき昆虫の羽根における翼間相互作用の空力的利点はストローク軌道に依存する」. Journal of Experimental Biology . 210 (Pt 8): 1362– 1377. doi : 10.1242/jeb.02746 . PMID 17401119. S2CID 23330782.
- ^ abcd Cheng, Xin; Sun, Mao (2016-05-11). 「ホバリング飛行中の小型昆虫の翼運動学測定と空気力学」. Scientific Reports . 6 (1) 25706. Bibcode :2016NatSR...625706C. doi :10.1038/srep25706. PMC 4863373. PMID 27168523 .
- ^ Kasoju, V.; Santhanakrishnan, A. (2021). 「翼型翼のフリングにおける空力相互作用」. Physics of Fluids . 33 (3): 031901. arXiv : 2011.00939 . Bibcode :2021PhFl...33c1901K. doi :10.1063/5.0036018. S2CID 226227261.
- ^ Weisberger, Mindy (2016年2月19日). 「蝶のように泳ぐ? カタツムリは水中を『飛ぶ』」. Scientific American . 2016年2月20日閲覧。
- ^ Murphy, D.; Adhikari, D.; Webster, D.; Yen, J. (2016). 「プランクトン性の海蝶による水中飛行」. Journal of Experimental Biology . 219 (4): 535– 543. doi : 10.1242/jeb.129205 . PMID 26889002.
- ^ 「昆虫の羽根一般」昆虫の空気力学、Cislunar Aerospace、1997年。 2011年3月28日閲覧。
- ^ 「シロチョウ科の蝶(白蝶)」Bumblebee.org . 2018年3月18日閲覧。
- ^ ab ダドリー、ロバート (2000). 『昆虫の飛翔のバイオメカニクス:形態、機能、進化』 プリンストン、ニュージャージー州:プリンストン大学出版局. p. 69. JSTOR j.ctv301g2x.
- ^ ab Stocks, Ian. (2008).翼結合に関する章、Capinera (Ed) (2008)「Encyc. Entom.」第4巻、p. 4266
- ^ スコブル、マルコム・J. (1995). 鱗翅目昆虫:形態、機能、多様性.自然史博物館. pp. 56– 60. ISBN 978-0-19-854952-9。
- ^ ab Powell, Jerry A. Ch.鱗翅目(pp. 631–664)Resh & Cardé(編)『昆虫百科事典』2003年。
- ^ Gorb, S. (2001) Ch 4.1.5「身体部位の連動」pp 46–50。
- ^ Woiwod, IP; Reynolds, DR; Thomas, CD(編)2001.「昆虫の移動:メカニズムと結果」CABインターナショナル。
- ^ Joos, B. (1987). 「 Manduca sextaの飛翔筋における飛行前ウォームアップ中の炭水化物利用」. Journal of Experimental Biology . 133 : 317–327 . doi :10.1242/jeb.133.1.317.
- ^ Sanjay P. Sane、Alexandre Dieudonné、Mark A. Willis、Thomas L. Daniel. 「蛾の飛行制御を媒介する触角機械センサー」 Science 315, no. 5813 (2007): 863-866.
- ^ ディッカーソン、ブラッドリー・H、アリシャ・M・デ・ソウザ、アイヌル・フーダ、マイケル・H・ディキンソン。「ハエは二重機能ジャイロスコープの能動制御によって羽の動きを制御する。」カレントバイオロジー29巻20号(2019年):3517-3524。
- ^ Wolf, Harald. 「イナゴのテグラ:飛行リズムの生成、羽ばたき運動の制御、そして空気力学的力の発揮における意義」Journal of Experimental Biology 182, no. 1 (1993): 229-253.
- ^ Misof, B.; Liu, S.; Meusemann, K.; Peters, RS; Donath, A.; et al. (2014). 「系統ゲノム解析は昆虫進化のタイミングとパターンを解明する」. Science . 346 (6210): 763– 767. Bibcode :2014Sci...346..763M. doi :10.1126/science.1257570. PMID 25378627. S2CID 36008925.
- ^ ブラウクマン、カールステン;シュナイダー、イェルク (1996)。 「Ein unter-karbonisches Insekt aus dem Raum Bitterfeld/Delitzsch (Pterygota, Arnsbergium, Deutschland)」[ビッターフェルト/デーリッチュ地域 (ドイツ、アルンスベルギアン、プテリゴタ) 産の下部石炭紀の昆虫]。Neues Jahrbuch für Geologie und Paläontologie - Monatshefte (ドイツ語)。1996 (1): 17–30。土井:10.1127/njgpm/1996/1996/17。
- ^ Tillyard, RJ (2009). 「ライニーチャート層(オールドレッドサンドストーン)産のデボン紀の化石昆虫に関する考察」.ロンドン王立昆虫学会誌. 76 (1): 65– 71. doi :10.1111/j.1365-2311.1928.tb01188.x.
- ^ ハウグ、カロリン;ハウグ、ヨアヒム (2017). 「推定最古の飛翔昆虫:多足動物の可能性が高い?」PeerJ . 5 e3402. doi : 10.7717/peerj.3402 . PMC 5452959. PMID 28584727 .
- ^ Wipfler, Benjamin; et al. (2019年2月19日). 「ポリ新翅目の進化史と初期の有翅昆虫の理解への示唆」. Proceedings of the National Academy of Sciences . 116 (8): 3024– 3029. Bibcode :2019PNAS..116.3024W. doi : 10.1073/pnas.1817794116 . PMC 6386694. PMID 30642969 .
- ^ Yanoviak, Stephen P.; Dudley, Robert (2018年7月). 「水生カゲロウ幼虫(Myobaetis ellenae, Baetidae)の跳躍と空中行動」.節足動物の構造と発達. 47 (4): 370– 374. Bibcode :2018ArtSD..47..370Y. doi : 10.1016/j.asd.2017.06.005 . PMID 28684306. S2CID 205697025.
- ^ ab Trueman, JWH (1990)、「コメント:昆虫の羽の進化:肢体退縮モデル」、Canadian Journal of Zoology。
- ^ ab Crampton, G. (1916). 「パラノータル説による昆虫の系統学的起源と羽の性質」ニューヨーク昆虫学会誌. 24 (1): 1– 39. JSTOR 25003692.
- ^ abc グリマルディ, デイヴィッド; エンゲル, マイケル・S. (2005). 『昆虫は空を飛ぶ』 ニューヨーク:ケンブリッジ大学出版局. pp. 155– 159.
{{cite book}}:|work=無視されました (ヘルプ) - ^ フリッツ、ミュラー(1875)イエナ。 Zeitung Naturwissenschaften、9、241。
- ^ Kukalova-Peck, Jarmila (1978). 「化石記録に見る昆虫の羽の起源と進化、そして変態との関係」. Journal of Morphology . 156 (1). Wiley: 53–125 . doi :10.1002/jmor.1051560104. PMID 30231597. S2CID 52301138.
- ^ Rasnitsyn, Alexander P. (1981). 「昆虫の翅の起源に関する修正された傍胸膜理論」. Journal of Morphology . 168 (3): 331– 338. doi :10.1002/jmor.1051680309. PMID 30110990. S2CID 52010764.
- ^ Thomas, Adrian LR ; Norberg, R. Åke (1996). 「表面をかすめる ― 昆虫の飛翔の起源?」Trends in Ecology & Evolution . 11 (5): 187– 188. Bibcode :1996TEcoE..11..187T. doi :10.1016/0169-5347(96)30022-0. PMID 21237803.
- ^ マーデン、ジェームズ (2003). 「昆虫の飛翔進化における表面スキミング仮説」. Acta Zoologica Cracoviensia 46 : 73–84 .
- ^ Yanoviak, Stephen P.; Kaspari, M.; Dudley, R. (2009). 「滑空する六脚動物と昆虫の空中行動の起源」. Biology Letters . 5 (4): 510–2 . doi :10.1098/rsbl.2009.0029. PMC 2781901. PMID 19324632 .
- ^ Averof, Michalis; Cohen, Stephen M. (1997). 「昆虫の羽の進化的起源:先祖の鰓から」Nature 385 ( 6617). Springer Science and Business Media: 627– 630. Bibcode :1997Natur.385..627A. doi :10.1038/385627a0. PMID 9024659. S2CID 4257270.
- ^ 丹羽尚; 秋元(加藤)愛; 新見輝之; 東條幸治; 町田隆一郎; 林茂雄 (2010-03-17). 「2つの発生モジュールの統合による昆虫の翅の進化的起源」.進化と発達. 12 (2): 168– 176. doi :10.1111/j.1525-142x.2010.00402.x. PMID 20433457. S2CID 15838166.
- ^ Elias-Neto, Moysés; Belles, Xavier (2016). 「背部および胸膜構造はゴキブリの異所性前胸翅形成に寄与する」. Royal Society Open Science . 3 (8) 160347. The Royal Society. Bibcode :2016RSOS....360347E. doi :10.1098/rsos.160347. PMC 5108966. PMID 27853616 .
- ^ 友安 良則; 大出 貴弘; クラーク・ハクテル コートニー (2017-03-14). 「連続ホモログからわかる昆虫の羽の起源」F1000Research . 6 : 268. doi : 10.12688/f1000research.10285.1 . PMC 5357031 . PMID 28357056.
- ^ プロコップ、ヤクブ;ペチャロバ、マルティナ。ネル、アンドレ。ヘルンシェマイヤー、トーマス;クシェミンスカ、エヴァ。クシェミンスキ、ヴィエスワフ。エンゲル、マイケル S. (2017)。 「古生代の若虫の羽パッドは昆虫の羽の起源のデュアルモデルをサポートします」。現在の生物学。27 (2): 263–269。書誌コード:2017CBio...27..263P。土井:10.1016/j.cub.2016.11.021。PMID 28089512。
さらに読む
- ディキンソン, MH; レーマン, FO; セイン, SP (1999). 「昆虫の飛翔における翼の回転と空気力学的基礎」. Science . 284 (5422): 1954–60 . doi :10.1126/science.284.5422.1954. PMID 10373107.
- エリントン, チャールズ・P.; ヴァン・デン・バーグ, コーエン; ウィルモット, アレクサンダー・P.;トーマス, エイドリアン・LR (1996). 「昆虫飛行における前縁渦」. Nature . 384 (6610): 626– 630. Bibcode :1996Natur.384..626E. doi :10.1038/384626a0. S2CID 4358428.
- エリントン, CP (1984). 「ホバリング昆虫飛行の空気力学 I. 準定常解析」.王立協会哲学論文集 B : 生物科学. 305 (1122): 1– 15.書誌コード:1984RSPTB.305....1E. doi :10.1098/rstb.1984.0049.
- エリントン, CP (1999). 「昆虫飛行の新しい空気力学:マイクロエアビークルへの応用」 .実験生物学ジャーナル. 202 (Pt 23): 3439– 3448. doi :10.1242/jeb.202.23.3439. PMID 10562527.
- Lewin, GC; Haj-Hariri, H. (2003). 「粘性流中における2次元ヒービング翼の推力発生モデル化」. Journal of Fluid Mechanics . 492 : 339–362 . Bibcode :2003JFM...492..339L. doi :10.1017/S0022112003005743. S2CID 122077834.
- プリングル、JWS (1975).昆虫の飛翔. オックスフォード生物学リーダーズ. 第52巻.
- Sane, SP (2003). 「昆虫の飛行における空気力学」. The Journal of Experimental Biology . 206 (Pt 23): 4191– 208. doi :10.1242/jeb.00663. PMID 14581590. S2CID 17453426.
- サベージ, SB; ニューマン, BG; ウォン, DTM (1979). 「トンボのホバリング飛行における渦と非定常効果の役割」 .実験生物学ジャーナル. 83 (1): 59– 77. doi :10.1242/jeb.83.1.59.
- Smyth, T. Jr. (1985). 「筋肉系」. Blum, MS. Blum (編).昆虫生理学の基礎. John Wiley and Sons . pp. 227– 286.
- Sotavalta, O. (1953). 「一部のユスリカにおける高頻度の羽ばたきと胸郭振動の記録」ウッズホール生物学紀要. 104 (3): 439– 444. doi :10.2307/1538496. JSTOR 1538496.
- Van Den Berg, C.; Ellington, CP (1997). 「ホバリングするスズメガの模型の渦後流」. Philosophical Transactions of the Royal Society B. 352 ( 1351): 317– 328. Bibcode :1997RSPTB.352..317V. doi :10.1098/rstb.1997.0023. PMC 1691928 .
- Walker, JA (2002年9月). 「回転揚力:何か違うのか、それとも同じことの繰り返しなのか?」 . The Journal of Experimental Biology . 205 (Pt 24): 3783– 3792. doi :10.1242/jeb.205.24.3783. PMID 12432002.
- Zbikowski, R. (2002). 「小型航空機におけるホバリング中の昆虫型羽ばたき翼の空力モデリングについて」. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences . 360 (1791): 273– 290. Bibcode :2002RSPTA.360..273Z. doi :10.1098/rsta.2001.0930. PMID: 16210181. S2CID : 2430367.
外部リンク
{kind=link}
- 小型航空機の開発における昆虫の役割
- 昆虫のような羽ばたき翼を持つ小型航空機
- 昆虫飛行の新しい空気力学:マイクロ航空機への応用
- 昆虫の飛行の空気力学
- 飛翔筋 アーカイブ 2004-08-22 at the Wayback Machine
- 蝶の空力機構の流れの可視化
- マルハナバチの飛行
- 昆虫の飛行